Joshua J. Schwartz – jschwartz2@pace.edu
Department of Biological Sciences
Pace University
Pleasantville, NY 10570
Popular version of paper 2pAB10 presented at the First Pan-American/Iberian Meeting on Acoustics, Cancun, Mexico
Regardless of what approach a female is using, the process will take time. If potential mates individually are inconsistent in attributes used by females or information about males is degraded as it travels through the environment to the female, increasing the amount of time devoted to mate assessment could help females more accurately gauge the quality of males. Relatively little information is available on just how long females engage in this assessment process. Here I describe how I obtained an estimate of the period of time that females of the gray treefrog allocate to assessment of the calling performance of nearby males. Specifically, this period (the assessment time) is the portion of time in which information gathered actually effects the female’s mate choice decision. Therefore it is different from search time, the period, including assessment time, during which a female hunts for an acceptable mate. Good estimates of this assessment period are not available for any species of frog in the wild.
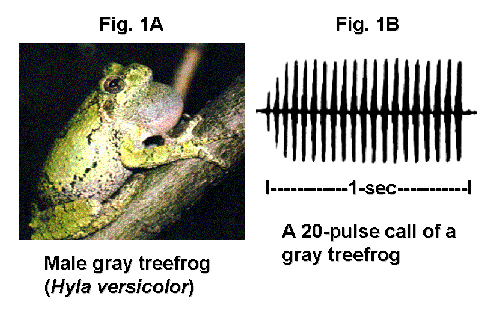
Males of the gray treefrog (Fig. 1A) call from vegetation in and adjacent to ponds during the late spring and summer in the Northeastern U.S. from dusk until near midnight. Females approach choruses from surrounding forest and can sit within a few feet of groups of males for more than one hour before they move toward a male and so solicit a reproductive coupling or ‘amplexus’. An extensive body of work by Dr. Carl Gerhardt (of the University of Missouri, Columbia) and his colleagues has demonstrated that females use the rate of calling by males as well as the duration of calls to discriminate among potential mates. Call duration, determined by the number of pulses in a call (Fig. 1B; click here to play a call), is particularly important in this regard. In fact, females will selectively discriminate against louder short calls in favor of less intense long calls, perhaps because genetic benefits accrue to the offspring of females that select males giving long calls. Males are quite sensitive to the amount of vocal activity at the pond and will add pulses to their calls in response to the calls of neighbors, presumably because this enhances their attractiveness under more competitive circumstances.
The majority of experiments designed to determine those call features female frogs prefer have been conducted in laboratory settings. Females that have been captured in amplexus are typically offered a choice between alternative calls broadcast from speakers located in a chamber lined with acoustic foam. Although this approach has proven very effective, for my study I decided to work entirely in the field with free-ranging females because it is possible previous pairing, capture and manipulation of test subjects alters the time window during which females compare calls during laboratory-based tests. However, I could not simply observe the behavior of female frogs in nature to obtain my estimate of assessment time for a couple reasons. First, I could not be certain that at any given moment a female in the vicinity of a group of calling males was actually attending to and comparing their advertisement signals. Second, even if females that I located were assessing males, I could not know when this process had started. Therefore, I decided to perform a female choice test in the field using calls that I would broadcast from speakers.
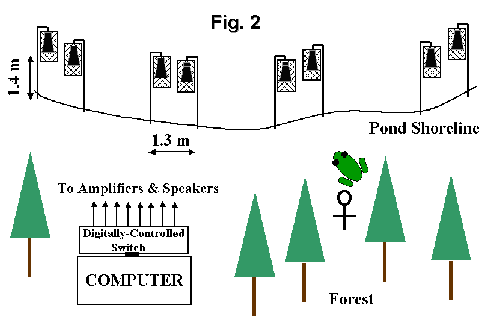
The experiment was conducted from mid-May to Mid-July at a pond in Ashland, Missouri. The frog calls were broadcast from eight outdoor patio speakers that radiated sound in a 360 pattern and were grouped in pairs at different points along the shore (Fig. 2). Each speaker was housed within a screened cage. Any females that might be attracted to the broadcast calls as they approached the artificial chorus from the adjacent forest gained access to a speaker through one of four vertical slits where the plastic mesh was folded and tied inward.
Each evening, my assistants and I hauled a computer and battery packs to the field site and mounted the speakers that had been stored in a metal bin from the previous night. It usually took us about 40 minutes to set up our equipment, adjust sound levels, and remove any males that were calling near the speakers. We usually began the playbacks by around 8:00 PM and ended them close to midnight. The calls were computer-synthesized versions of the natural call and were routed to the eight-speakers using hardware and software created for the experiment. One speaker in each speaker-pair always broadcast calls with the same number of pulses (12). The second speaker alternated between runs of very short calls (6 pulses) and runs of long calls (32 pulses). However, the amount of time devoted to these bouts of 6 pulse and 32 pulse calls differed at the four spots at the pond where this cycling stimulus was broadcast. These times were 2 minutes, 4 minutes, 16 minutes, and 32 minutes.
What was the rationale for using these particular calls and patterns of calling? Results from laboratory experiments indicated that female gray treefrogs should choose 36 pulse calls over 12 pulse calls and 12 pulse calls over 6 pulse calls. Laboratory tests also demonstrated that females are sensitive to subtle differences in the total number of pulses from speakers broadcasting calls with variable pulse-numbers and non-variable pulse-numbers. In the field experiment, females attracted to the calls coming from a particular pair of speakers would sometimes hear the very short calls offered against the calls of intermediate duration. Other times they would hear the long calls offered against the intermediate duration calls. If the time period over which females assess calling is so brief that they were only faced with one of these two situations, one would expect a 50 – 50 split in the number of females captured in the cages surrounding the speaker broadcasting the cycling stimulus and the cages surrounding the constant duration stimulus. However, with progressively longer periods of assessment, it was more likely that females would have heard both long and short calls broadcast from the cycling speaker. Thus they were increasingly likely to approach the cycling speaker as the discrepancy between the total number of pulses from the constant and cycling speakers increased within the assessment time window. If the period of assessment were long enough (more than 65 % of the time between successive broadcasts of the same call type from the cycling speaker), all females should have chosen the cycling speaker.
The actual number of females captured at each speaker of a particular pair of speakers provided an estimate of how long females had actually assessed the calling of the simulated males. Accordingly, at the end of our field season our capture results at the four locations at the pond gave us four independent estimates of the assessment period. Using a statistical approach that combined the results from all the speakers, it was possible to estimate the most likely time period of female assessment, 144 seconds. This result does not mean that variation does not exist among and within females in their assessment time. Obtaining the estimate relied upon the number of captures at different stimulus treatments and so details pertaining to individual differences could not be detected. Therefore, the result should be interpreted to mean that females choose a mate based on calling behavior occurring over minutes or less rather than tens of minutes. The estimate also does not mean that females do not listen to males for many minutes before selecting a mate. Rather, it means that females appear to use only the information in the calls given in about two minutes prior to pairing to make their choice of a mate. Patterns of variation in call duration both within and among different males observed in other experiments suggest that this assessment period is sufficient to allow females to distinguish among males in a way that accurately reflects male performance over longer blocks of time.
Work supported by NSF Grant IBN-9727623